An apparent direct negative relationship between large granules and starch digestibility was reported in 1922. Many studies have since confirmed this relationship. Lindeboom reported that small barley and wheat starch granules are hydrolyzed faster than large granules. Singh et al. observed significant differences between the enzymatic hydrolysis of different native potato starches when fractionated according to small, medium, and large granules.
- Introduction to Slowly Digestible Starch (SDS)
- Preparation of Slowly Digestible Starch (SDS)
- Physicochemical Properties of Slowly Digestible Starch (SDS)
- Functions of Slowly Digestible Starch (SDS)
- Applications of Slowly Digestible Starch (SDS)
The lower susceptibility of large granule starches to enzymatic hydrolysis has been attributed to the smaller specific surface area of the granules, which may reduce the extent of enzyme binding and ultimately result in less hydrolysis that occurs in small granules.
How the structural aspects of Digestible Starch (SDS) are related to the mechanisms of its digestion and their health implications have been investigated and reported. Based on current data, the slow digestibility of SDS is caused by two factors, its physical structure and chemical construction, both of which reduce the contact between enzymes and SDS.
Physical Structure of Slowly Digestible Starch (SDS) and Its Digestibility
The digestion of starch granules is a complex process that includes different phases: the diffusion of the enzyme toward the substrate, which is affected by the porosity of the substrate; the adsorption of the enzyme to the starchy material; and the hydrolytic event. A large amount of starch is protected from the protein and other components of food, and is not contacted by digestive enzymes, which is why pasta contains so much Digestible Starch (SDS).
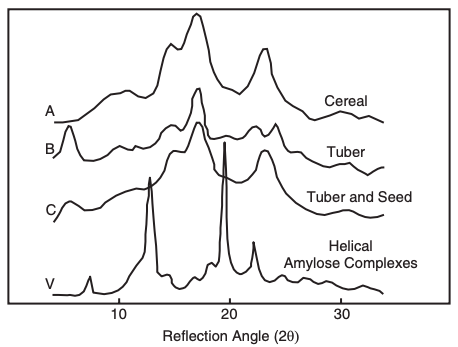
in starch
X-ray diffraction scattering studies have classified native starches into types A, B, C, and V (Figure 1). The molecular structure of the starch granules, especially the arrangements in A-type and B-type crystallites, influences the hydrolysis of the starches. A-type polymers are less resistant to amylase hydrolysis than B-type polymers. The shorter double helices and interior crystallites present in A-type starches are more susceptible, whereas long chains form longer and more stable helices and are more resistant to enzymatic hydrolysis. The structural arrangements of the A-type or B-type crystallites markedly influence their digestibility. Generally, a higher susceptibility to hydrolysis has been reported for the A-type crystallites than the B-type crystallites. A-type and B-type starches differ in the packing of their double helices and in their water content. The shorter double helices and interior crystallites of the A-type starches are more readily digestible and contain larger amounts of RDS and SDS than the B-type starches.
A comprehensive account of the enzymatic digestibility of native uncooked starches from different sources has been given by Dreher et al. The authors suggest that cereal starches are more digestible than tuber and legume starches, which may be attributable to the presence of numerous pinholes in the surface layers of the granules and pores, which penetrate toward the interior of the granules from cereal sources, such as corn. These pores in the granules facilitate the entry of amylases, allowing the digestion of the granules.

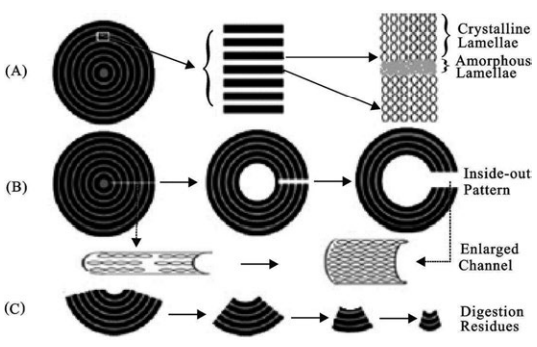
Huber et al. demonstrated that both the crystalline and amorphous regions in cereal starch granules have the same granule center, as shown in Figure 2. Enzymatic digestion begins at the surface pores and interior channels, and then digestion gradually enlarges the channel by simultaneously digesting the crystalline and amorphous regions. This explains this slow digestion process. Figure 3 shows that the digestion of the starch granules starts at the interior channels.
In general, starch is consumed after it is processed. An excess of water and high temperatures during processing cause starch gelatinization and destroy its granular structure. However, the granular structure of starch is retained in several lowmoisture food products, such as biscuits.
The hydrolysis of native starches can vary greatly, depending on the interplay of a range of factors, but it is usually determined by the botanical origin of the starch, which determines its morphology and crystalline organization. This offers a way to influence starch digestibility with breeding research and the selection of suitable crop varieties.
The diffusion of α-amylase into the substrate is considered an important step in hydrolysis. The interactions of starch with fiber, protein, and other food components can prevent the effective diffusion and adsorption of α-amylase. The hydrolysis of starch was previously considered to start from the surface of the granule. However, native cereal starches, such as corn and sorghum starches, contain peripheral pores and channels that allow the penetration of α-amylase, resulting in an inside-out hydrolysis mechanism. In contrast, potato starch and other B-type starches are digested from the surface of the starch granule, explaining the higher digestibility of cereal starches compared with tuber starches, such as potato starch. Tuber starches are generally more resistant to enzymatic hydrolysis than cereal starches, because of their larger granules, their surface properties, and their supramolecular arrangement. The large amounts of resistant starch in tubers, particularly the potato, and in fruits such as the banana have been reported both in vitro and in vivo, whereas cereal starches, such as rice, wheat, and barley starches, are highly digestible. Native normal cornstarch, waxy starches, millet starches, sorghum starches, and legume starch reportedly display intermediate digestibility because they contain medium-to-high amounts of SDS. Part of the lower digestibility of millet, sorghum, and legume starches can be attributed to their interactions with protein, which forms a protective network around the granule.
Fine Structure of Digestible Starch (SDS) and Its Digestibility
When starch is processed for use in food, the starch is gelatinized. At this time, the molecular structure of the starch is the only factor determining its functionality and nutritive peculiarities. The molecular structure of starch includes the ratio of amylose and amylopectin and the fine structure of amylopectin.
At the molecular level, the crystallite structure and the packing of the amorphous phase influence the enzymatic susceptibility of starch. The unit chain length of amylopectin correlates with its digestibility, and the proportion of the amylopectin unit chain length with DP 8–12 or DP 16–26 correlates positively and negatively, respectively, with its hydrolysis. Longer chains form longer and more stable helices, which are further stabilized by hydrogen bonds distributed throughout the entire crystalline region, which further reduce its digestibility.
The nature of starch also influences its digestibility and the postprandial glucose response. Starches high in amylopectin have been shown to be digested more quickly than those high in amylose, probably because amylopectin has many more nonreducing chain ends than amylose, to which digestive enzymes can attach.
Some researchers have shown that the fine structure of amylopectin, with its high branching density and either long or short internal chains, causes its slow digestion. Therefore, the inherent molecular structure of amylopectin is responsible for its slow digestion. The structure of Digestible Starch (SDS) may include imperfect crystallites and amylopectin, with a high branching density, which probably cause its slow digestion.
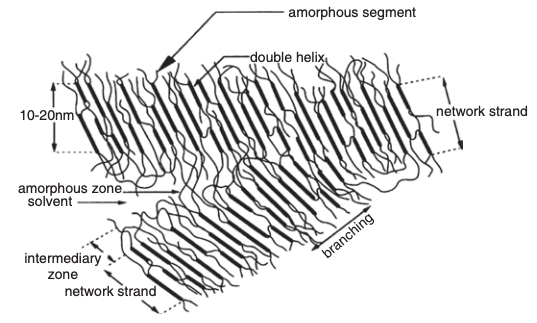
An illustration of retrograded starch chains and the arrangement of the amorphous material that determine the formation of SDS is presented in Figure 4. It is proposed that amylose chains aggregate in an infinite three-dimensional network, linking the microstructure and the macromolecular organization of starch. The chain segments inside the crystallites are disposed obliquely relative to the microfiber axis. Therefore, the network of strands consists of contiguous associated blocks aligned along the length axis of the microfiber. Double helices are then linked to other helices by loops of amorphous amylose segments, dangling in the gel pores. This fraction is responsible for the hydrodynamic behavior of amylose gels.