The size of amylose molecules is more frequently given as the DP than as molecular weight and is obtained by light scattering (Hizukuri and Takagi, 1984), from the limiting viscosity number (Cowie and Greenwood, 1957), or by reducing end analysis with the ParkeJohnson reagent (Park and Johnson, 1949) as modified by Hizukuri et al. (Hizukuri et al., 1981, 1983). The number-average DP (DPn) from different plants varies between 0.51 and 6.34X103 (Hizukuri et al., 1981; Takeda et al., 1984, 1986, 1988; Shibanuma et al., 1994; Schulman et al., 1995; Hizukuri and Takagi, 1984; Biliaderis et al., 1981; Lii and Chang, 1991). Weight-average values are higher (Takeda et al., 1986, 1999; Hizukuri and Takagi, 1984). The size distribution was analyzed by HPSEC on Toyosoda TSK-GEL PW columns, coupled to simultaneous refractive index (RI) and low-angle laser light scattering (LALLS) detection, for a range of samples (Hizukuri and Takagi, 1984; Takagi and Hizukuri, 1984). Broad distributions were obtained for amylose from potato (DPw 0.84-21.8X103) and tapioca (0.58-22.4X103), whereas amylose from kudzu possessed a more narrow distribution (0.48-12.3X103). More recently, multi-angle light scattering (MALS) detectors have largely replaced LALLS for molecular weight estimations (Ong et al., 1994; You and Lim, 2000; You and Izydorczyk, 2002; Zhong et al., 2006; Chen and Bergman, 2007).
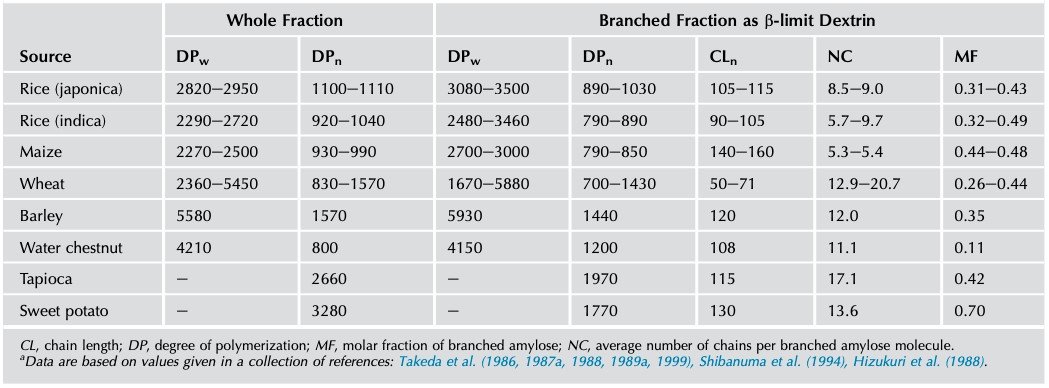
A considerable portion of the amylose fraction, between 10% and 70% depending on the sample, contains slightly branched macromolecules (Hizukuri et al., 1981; Manners and Bathgate, 1969; Takeda et al., 1987a). These are generally larger than the linear amyloses (Takeda et al., 1992b, 1993a) and possess between 5 and 20 chains per molecule. The C-chain of branched amylose from maize was tritium labeled at the reducing end by treatment with sodium [3H]-borohydride and CL ranged from 200 to 710 (Takeda et al., 1992a). Takeda et al. (1987a, 1989a, 1992b) studied branched amyloses by preparing the β-amylolysis limit dextrins (β-LDs) of the amylose fraction. All linear amylose, together with the external chains of the branched amylose, is hydrolyzed. The molar fraction of branched amylose (MFbranched) is calculated from (Shibanuma et al., 1994; Takeda et al., 1987a):
MFbranched = (NCwhole amylose sample/(NCβ-LD-1)
in which NC is the average number of chains per molecule.
Takeda et al. (1990) subfractionated maize amylose in aqueous n-butanol. A minor, branched component with high molecular weight remained in the supernatant. It was shown that the component contained a fraction of very short chains that were suggested to form small “immature” clusters. A somewhat similar conclusion was made by Takeda et al. (1993a) for rice amylose. After debranching the rice amylose (with the enzyme isoamylase), these authors labeled the unit chains with tritium and found that the amylose not only contained long chains typical of amylose but also short chains possessing a peak at DP 21 when analyzed by GPC. In fact, these short chains had lengths comparable to chains typically found in amylopectin. Moreover, on a number basis, the short chains predominated: the molar ratio of very long (DP > 200), long (DP 30-200), and short chains (DP 10-30) was 0.8:0.6:4.0. Hanahiro et al. (2013) confirmed the existence of short chains in rice amylose using fluorescent labeling with 2-aminopyridine. Based on several details in the size-distribution pattern of the chains, Hanahiro et al. (2013) concluded that the branching pattern of these short chains in amylose was distinct from the branching pattern of amylopectin.